
Botanical origin and biodiversity of maize: consequences for its productivity and nutritional value
The amazing modifications which led to the maize plant determined its world success. Nevertheless, those modifications are so important that for a long time we no longer could identify the wild ancestor of maize. On the other hand, those modifications had major consequences on the future of this plant.
La domestication de Zea mays Parviglumis n'aurait modifié qu'une dizaine de gènes majeurs selon les généticiens. Particulièrement tb1 (branches et position des épis), tga1 (enveloppe dure du grain).
La première mutation sélectionnée par l'homme est probablement celle qui fait que le grain, à maturité, reste solidement attaché sur un épi lui-même rigide. En cherchant, dans la descendance d'un croisement maïs / téosinte, les plantes les plus proches de la téosinte, on trouve de petits épis à deux rangs.

L'homme a réussi cette transformation sur toutes les plantes cultivées.
Elle permet une récolte beaucoup plus rapide et sûre mais rend toutes les plantes cultivées (pas seulement le maïs comme on le lit parfois) incapables de se disséminer. Elle devront passer obligatoirement par deux opérations humaines : égrenage et semis pour se reproduire.
L'homme a donc gardé une mutation que la nature n'aurait et n'a jamais gardée car elle condamnerait l'espèce sauvage.
 sont entourés d'une coque dure, ici ouverte, alors que les grains de maïs sont nus.") La deuxième mutation est celle qui supprime la coque dure qui protège la graine de téosinte. Le maïs a lui un « grain nu » (comme le blé), une autre transformation anti-naturelle qui aboutit à exposer la graine aux insectes et aux champignons mais qui la rend infiniment plus digestible pour l'homme et ses animaux domestiques monogastriques, porcs et surtout volailles, sans avoir a pratiquer un décorticage long et énergivore.
La deuxième mutation est celle qui supprime la coque dure qui protège la graine de téosinte. Le maïs a lui un « grain nu » (comme le blé), une autre transformation anti-naturelle qui aboutit à exposer la graine aux insectes et aux champignons mais qui la rend infiniment plus digestible pour l'homme et ses animaux domestiques monogastriques, porcs et surtout volailles, sans avoir a pratiquer un décorticage long et énergivore.
Le succès actuel du blé et du maïs sur le grand marché mondial de l'alimentation animale est en grande partie dû à cette modification génétique millénaire !
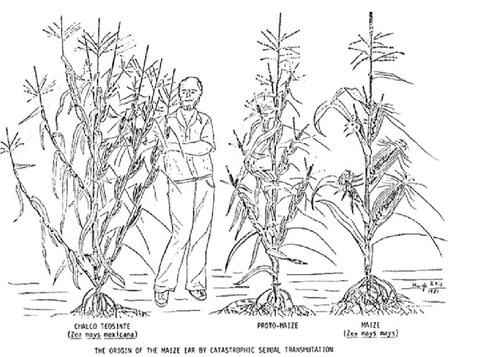 L'architecture de la plante est aussi totalement modifiée. Le nombre de branches latérales de la téosinte est considérablement réduit et les deux restantes (en général) sont raccourcies au point de couvrir avec leurs feuilles l'épi de maïs qui est une évolution de la panicule de la branche latérale, transformée en fleur femelle !
L'architecture de la plante est aussi totalement modifiée. Le nombre de branches latérales de la téosinte est considérablement réduit et les deux restantes (en général) sont raccourcies au point de couvrir avec leurs feuilles l'épi de maïs qui est une évolution de la panicule de la branche latérale, transformée en fleur femelle !
Incroyable tour de force dont le maïs a l'exclusivité et qui permet de situer la masse de grain à mi-hauteur de la tige (au lieu de l'extrémité comme dans toutes les graminées domestiques) et d'augmenter ainsi la capacité de la plante à supporter de gros rendement en grain sans verser.
Le coup de génie final des premiers agriculteurs Amérindiens fut de recouvrir et protéger les grains nus par les feuilles de l'ancienne branche latérale. On les appelle maintenant spathes... et une opération humaine de plus pour reproduire le maïs : l'effeuillage qui justifia nos belle fêtes nocturnes Pyrénéennes des « espelouquères ».
 La taille du grain, enfin, a été considérablement augmentée, facilitant égrenage et mouture et réduisant le déjà faible taux de membranes cellulosiques indigestes pour les animaux monogastriques, homme inclus.
La taille du grain, enfin, a été considérablement augmentée, facilitant égrenage et mouture et réduisant le déjà faible taux de membranes cellulosiques indigestes pour les animaux monogastriques, homme inclus.
 Les étapes suivantes, qui ont duré des millénaires, ont abouti à une diversification étonnante des couleurs et types de grains, depuis les « pop corns » aux minuscules grains totalement vitreux, jusqu'aux maïs farineux et aux maïs doux riches en sucre.
Les étapes suivantes, qui ont duré des millénaires, ont abouti à une diversification étonnante des couleurs et types de grains, depuis les « pop corns » aux minuscules grains totalement vitreux, jusqu'aux maïs farineux et aux maïs doux riches en sucre.
 En Europe nous classons nos maïs en : grains vitreux, cornés (majorité d'amidon vitreux), cornés dentés, dentés (majorité amidon farineux).
En Europe nous classons nos maïs en : grains vitreux, cornés (majorité d'amidon vitreux), cornés dentés, dentés (majorité amidon farineux).
Toutes ces diversifications étaient déjà accomplies lorsque les européens découvrirent l'Amérique. D'autres caractéristiques de la téosinte sont bien sûr restées intactes et caractérisent toujours le maïs.
Photosynthèse en C4
Comme d'autres graminées tropicales, au cours de l'évolution (Canne à sucre, sorgho, mil, miscanthus), la téosinte et donc le maïs sont des plantes « en C4 ». Ce terme vient d'une particularité de leur photosynthèse qui concentre le carbone en composant à 4 atomes, acides organiques ou leurs sels (oxaloacétate, malate, aspartate).
Ce processus métabolique intervient préalablement au cycle de Calvin, mais dans un tissu différent, augmentant le rendement de la photosynthèse : une sorte de « turbo ». Les plantes en C4 produisent de l'ordre de 40 kg de matière sèche par ha/mm d'eau alors que les autres, en C3, n'en produisent que 30.
Si on considère uniquement l'aspect efficience de l'utilisation de l'eau, on observe que pour produire 1 kg de matière sèche (= environ 0,5 kg de grain) le maïs a besoin de 450 l d'eau, le blé de 590, le soja de 900, le riz plus de 5 000 (chiffres CNRS rapportés par AGPM).
La supériorité des plantes en C4 est aussi manifeste en conditions chaudes (pour un même rendement que les C3 , les stomates sont plus fermés, donc moins de déperdition d'eau) et au niveau de l'efficience d'utilisation de l'azote.
Système racinaire
 Il est superficiel, caractéristique des plantes croissant en conditions humides : racines relativement fines et réparties à 80 % dans les premiers 20 cm du sol. 10 000 ans de sélection n'ont pas modifié ce caractère qui explique les besoins d'une irrigation de qualité en zones à été sec.
Il est superficiel, caractéristique des plantes croissant en conditions humides : racines relativement fines et réparties à 80 % dans les premiers 20 cm du sol. 10 000 ans de sélection n'ont pas modifié ce caractère qui explique les besoins d'une irrigation de qualité en zones à été sec.
Le mode de reproduction a été probablement partiellement modifié par la nouvelle architecture de la plante.
La monoecie et l'allogamie anémophile viennent bien de la téosinte, la dichogamie a peut être été induite par 9 000 ans de sélection massale et reste d'ailleurs variable.
La monoecie signifie que les fleurs femelles et mâle se trouvent sur la même plante et elle sont séparées sur la plante chez le maïs comme chez la téosinte. Ce caractère facilite actuellement la fabrication d'hybrides commerciaux au champ.
L'allogamie anémophile (fécondation croisée et pollen transporté par le vent) explique le caractère hybride très majoritaire de la reproduction du maïs. Une population (variété de pays en fécondation libre) produit 89 à 95 % de grains hybrides (W. Wouters 1941) et les quelques auto-fécondations sont éliminées par les pratiques d'éclaircissage et de sélection massale.
Une variété ancienne est aussi hybride qu'un hybride F1 moderne. D'une façon générale la reproduction hybride du maïs a toujours facilité le maintient de sa biodiversité allant même jusqu'à permettre sa recombinaison spontanée avec son ancêtre téosinte au Mexique.
La dichogamie (décalage de floraison mâle et femelle) favorise aussi l'hybridation, elle est très majoritairement protandre (mâle en premier) mais les efforts récents des sélectionneurs pour réduire l'ASI (Intervalle anthères-soies) afin d' améliorer la tolérance à la sècheresse, a produit des variétés plus synchrones voire protogynes.
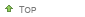

Épi et graines de téosinte